研究内容
[4] 高圧増殖に関与する新規ERタンパク質Ehg1の役割
ゲノムレベルで見た高圧・低温条件下の増殖に必要な遺伝子
出芽酵母には6,000個以上の遺伝子が存在しますが、うち約4,800個は単独破壊しても致死とはなりません。私たちはそれら遺伝子破壊株のライブラリーであるYeast deletionsを高圧・低温下でスクリーニングし、84個の感受性株を取得しました(Abe and Minegishi, Genetics 2008)。すなわち少なくとも84個の遺伝子が酵母の高圧・低温増殖に不可欠といえます(図1)。用いた親株は自らトリプトファンを合成できるため、高圧(25MPa, 25°C)や低温(0.1MPa, 15°C)で増殖することができます(ただし、ロイシン、ヒスチジン、リジンおよびウラシルを作れません)。予想通りトリプトファン合成系に失陥があると、酵母はこうした条件下で増殖できなくなります。また、新たにフェニルアラニンやチロシン、セリン、スレオニンといったアミノ酸の合成系にも重要性が見いだされました(図1)。それら破壊株の増殖は培養液中のアミノ酸に依存するので、裏を返すと上記アミノ酸の取り込みが高圧や低温下で阻害されることを意味しています。そこには12個の機能未知遺伝子が含まれていたのですが、最近、興味深い事実が明らかとなりました。
84個の遺伝子破壊株全てに対し栄養要求性を相補する4つのプラスミド(LEU2, HIS3, LYS2およびURA3)を同時に導入したところ、24個の破壊株において野生株レベルまで高圧増殖能が回復したのです。しかも顕著だったのが12未知遺伝子破壊株のうちの7株でした(Kurosaka et al., Sci. Rep. 2019)。つまりロイシン、ヒスチジン、リジンおよびウラシルを自ら合成できるようにすると高圧増殖能が回復するということは、破壊株ではこれら栄養源の取り込みが高圧下で損なわれていることを意味します。従ってこれら未知遺伝子の産物は、栄養源輸送体が高圧下で機能するために重要な役割を果たしているといえます。さらに興味深いことは、Costanzoらが行った遺伝子の二重破壊株の網羅的解析から、これら12未知遺伝子のうち6個が見事なクラスターを形成していたことです(Costanzo et al., Science 2016)。このことからも6個の機能未知タンパク質が連携して高圧下における生存を可能にしているのです。
YPR153W/MAY24は小胞体膜に局在するタンパク質をコードしていました。そこでこれをER-associated high-pressure growth gene 1 (EHG1)と名付けました(図2, Kurosaka et al., Sci. Rep. 2019)。Split ubiquitinシステムを利用した分子間相互作用解析から、Ehg1は栄養源輸送体と物理的に相互作用することもわかりました。そして、最近私たちは、Ehg1が表層小胞体(cER)で細胞膜(PM)上のトリプトファン輸送体Tat2と物理的に相互作用し、Tat2を”脂質ラフト”と呼ばれるドメイン構造に安定化する機能を持つことを突き止めました(図3, Kato et al. Mol. Biol. Cell 2025)。高圧や低温などのストレス条件にさらされると生体膜構造に歪みが進み、Tat2などのアミノ酸輸送体は不安定化します。Tat2はEhg1の働きによって脂質ラフトで安定化し、こうしたストレス下でも正常な機能を果たせるようになるのです。
 図1. 高圧・低温増殖に必須な遺伝子群 図1. 高圧・低温増殖に必須な遺伝子群 |  図2. Ehg1は新規ER膜タンパク質だった。 図2. Ehg1は新規ER膜タンパク質だった。 | 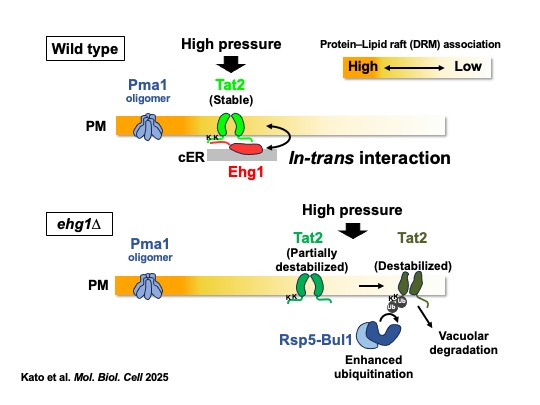 図3. Ehg1はアミノ酸輸送体の脂質ラフト局在を促進する。 図3. Ehg1はアミノ酸輸送体の脂質ラフト局在を促進する。 |